拓扑异构酶_拓扑异构酶的分类
拓扑异构酶(topoisomerase)是指通过切断DNA的一条或两条链中的磷酸二酯键,然后重新缠绕和封口来更正DNA连环数的酶。
拓扑异构酶(topoisomerase)是指通过切断DNA的一条或两条链中的磷酸二酯键,然后重新缠绕和封口来更正DNA连环数的酶。
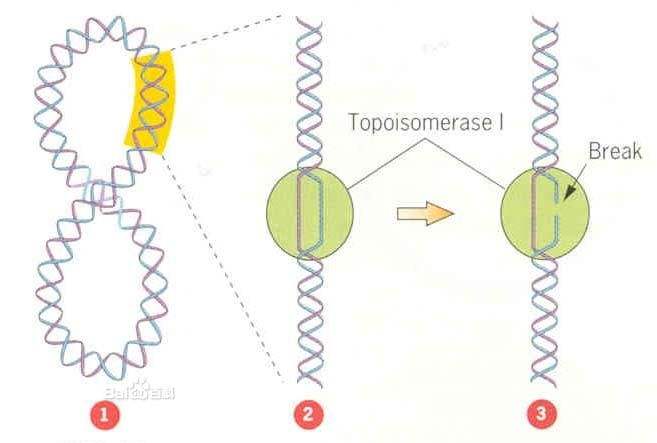
DNA拓扑异构酶是存在于细胞核内的一类酶,他们能够催化DNA链的断裂和结合,从而控制DNA的拓扑状态,拓扑异构酶参与了超螺旋结构模板的调节。哺乳动物中主要存在两种拓扑异构酶。
DNA拓扑异构酶I通过形成短暂的单链裂解-结合循环,催化DNA复制的拓扑异构状态的变化;相反,拓扑异构酶II通过引起瞬间双链酶桥的断裂,然后打通和再封闭,以改变DNA的拓扑状态。
哺乳动物拓扑异构酶II又可以分为αII型和βII型。拓扑异构酶毒素类药物的抗肿瘤活性与其对酶-DNA可分裂复合物的稳定性相关。这类药物通过稳定酶-DNA可分裂复合物,有效地将酶转换成纤维毒素。
拓扑异构酶可分为两类一类叫拓扑异构酶I,一类叫拓扑异构酶II。
1、DNA拓扑异构酶I
拓扑异构酶I催化DNA链的断裂和重新连接,每次只作用于一条链,即催化瞬时的单链的断裂和连接,它们不需要能量辅因子如ATP或NAD。E.coliDNA拓扑异构酶I又称ω蛋白,大白鼠肝DNA拓扑异构酶I又称切刻-封闭酶。
DNA拓扑异构酶能催化的反应很多,如DNA拓扑异构酶I对单链DNA的亲和力要比双链高得多,这正是它识别负超螺旋DNA的分子基础,因为负超螺旋DNA常常会有一定程度的单链区。
负超螺旋越高,DNA拓扑异构酶I作用越快。现已知道,生物体内负超螺旋稳定在5%25左右,低了不行,高了也不行。生物体通过拓扑异构酶I和II的相反作用而使负超螺旋达到一个稳定状态。
现已发现,编码E.coli拓扑异构酶I的基因topA发生突变,则会引起旋转酶基因的代偿性突变;否则,负超螺旋增高,细胞生活能力降低。
拓扑异构酶I作用的碱基序列特异性不高,但切点一定在C的下游方向4个碱基(包括C本身)的位置。在将DNA单链切断后,拓扑异构酶I连接于切口的5端,并贮藏了水解磷酸二脂键的能量用以连接切口,因而拓扑异构酶I的作用不需能量供应。
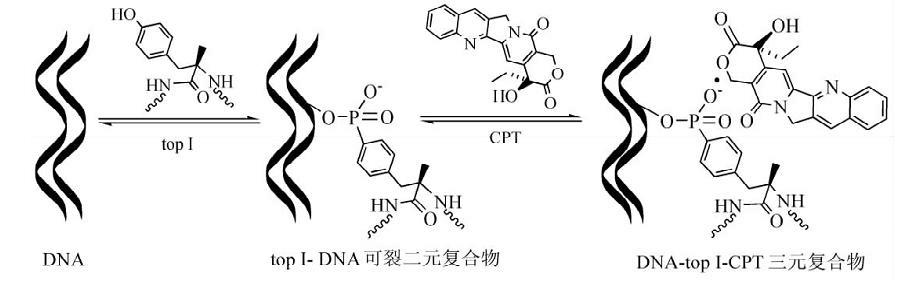
此外.拓扑异构酶I还能促进两个单链环的复性,其作用是解除复性过程所产生的链环数的负值压力,以使复性过程进行到底。
如果在一个单链环上一个部位切断,而使另一部位绕过切口.则可产生三叶形结构分子。如果有两个双链环,其中一个有一个切刻,拓扑异构酶I则可以将切刻对面的一条链切断,使完整的双链环套进去,再连接起来而成为环连体分子。
拓扑异构酶Ⅰ最早是1971年在大肠杆菌中被发现的,均为单体酶。拓扑异构酶Ⅰ根据其结构域功能可以划分为4个域C端结构域、核心结构域、连接子区域和N端域,其中C端结构域、核心结构域在催化活性中起主要作用。
在拓扑异构酶Ⅰ的多个活性位点中,Arg488、Arg590、His632和Tyr723为研究比较明确的4个活性位点。除Tyr723位于羧基端结构域外,其余三个均位于拓扑异构酶Ⅰ的核心结构域。
2、DNA拓扑异构酶II
拓扑异构酶II能同时断裂并连接双股DNA链.它们通常需要能量辅因子ATP。在拓扑异构酶II中又可以分为两个亚类:一个亚类是DNA旋转酶,其主要功能为引入负超螺旋,在DNA复制中起十分重要的作用。
迄今为止,只有在原核生物中才发现DNA旋转酶,另一个亚类是转变超螺旋DNA(包括正超螺旋和负超螺旋)成为没有超螺旋的松弛形式。
这一反应虽然是热力学上有利的方向,但不知道为什么它们仍然像DNA旋转酶一样需要ATP,这可能与恢复酶的构象有关。这一类酶在原核生物和真核生物中都有发现。
大肠杆菌的拓扑异构酶II除了引入负超螺旋以外.还具有形成或拆开双链DNA环连体和成结分子的能力。II类拓扑异构酶没有碱基序列特异性,它们可以和任何相交的两对双链DNA结合。
DNA旋转酶有两个α亚基和两个β亚基。α亚基约105KD,为gyrA基因所编码,具有磷酸二脂酶活性,可为萘啶酮酸所抑制。β亚基约95KD,为graB基因所编码,具有ATP酶活性,可为新生霉素所抑制。
这两种药物均可抑制野生型大肠杆菌的DNA复制。可见DNA旋转酶为E.coli的复制所不可缺少的。在切断一条DNA双链后,两个a亚基各结合于切口的一个5端,并贮藏了水解磷酸二酯键而获得的能量,由于该酶的整体性,因而DNA链的四个断头并无任意旋转的可能性。
由于酶的别构效应,使完整的双链穿过切口,然后再重新形成磷酸二酯键。β亚基的功能在于水解ATP以使酶分子恢复原来的构象,以便进行下一轮反应。
这一点可以用ATP的同系物β,γ-亚氨基ATP代替ATP而得到证实。因为这一同系物不能被DNA旋转酶所水解,但它确能促进第一轮拓扑异构反应,使负超螺旋增加,而妨碍以后进一步的拓扑异构反应。
真核生物拓扑异构酶Ⅱ为同源二聚体,包括的两个亚型拓扑异构酶Ⅱα和拓扑异构酶Ⅱβ分别定位于染色体17q21—22[10]和3p24[11]单拷贝基因编码的二聚体蛋白。
拓扑异构酶Ⅱ可划分为三个不同区域%3AC端域、N%20端域(ATP结合域)以及中部功能域(如下图右)。DNA拓扑异构酶的C端域在其对DNA的构象识别方面起主要作用,N端域和中部功能域则为拓扑异构酶的主要活性域。
以上就是关于拓扑异构酶分类的介绍,更多资讯请关注本站WAM机械网!